New paper / Dear Reviewer #2
While I was developing my E. coli reconstitution of a bacterial CO2 concentrating mechanism (CCM), I tested my rubisco-dependent E. coli strains in a variety of CO2 levels. As a reminder, CO2 is the rubisco carboxylation substrate. Normally we consider atmosphere (0.04% CO2) to be “low” and 5-10% CO2 to be “high,” but I wanted to try intermediate levels to understand what was going on. I noticed that several control strains had odd growth phenotypes in ≈1% CO2 atmosphere. In particular, strains that expressed bicarbonate (HCO3-) transporters but no other CCM components grew better at 1% CO2 than controls expressing only rubisco. This was super confusing because the rubisco substrate is clearly CO2 and not HCO3-. Working to understand this phenotype sent me and Eli on a multi-year adventure that introduced us to the Earth sciences and segued into my postdoc at Caltech via collaborating with Woody.
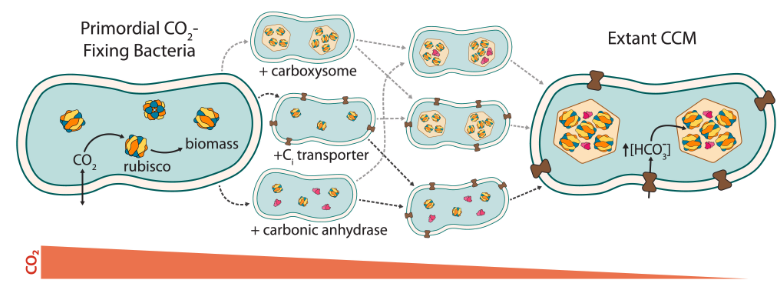
Our new paper - Trajectories for the evolution of bacterial CO2-concentrating mechanisms - aims to understand how CCMs evolved. Why is that even a question? Well, because all modern Cyanobacteria have CCMs composed of multiple interoperating protein complexes. Each complex is made of many protein subunits, and mutants lacking any core CCM gene are dead. That is, they totally fail to grow in today’s atmosphere (0.04% CO2). If any individual mutant is lethal, how could the CCM have evolved? Like the eye, it’s possible that ancestral CCMs had a slightly different function than modern forms. Just as ancient eyes sensed light rather than resolving images, ancient CCMs might have performed alternate roles. This hypothesis is bolstered by some evidence from Earth sciences, where several approaches to estimating the CO2 content of Earth’s ancient atmopshere indicate that historical CO2 levels were much greater 1-3 billion years ago than they are today. So perhaps ancient CCMs just had a simpler task?
To address this hypothesis - that the history of atmospheric CO2 can explain CCM evolution - Eli and I used the tools of synthetic biology. Using our E. coli CCM reconstitution we designed modern-day bacteria engineered to resemble potential CCM ancestors. We did this by removing some crucial functions, including breaking structures, enzymes and transporters in targetted ways. We then grew these “putative ancestral analogs” in a variety of CO2 concentrations to simulate Earth’s historical atmosphere.
Surprisingly, we found at least two “fitness-positive” paths for the “CO2-catalyzed” evolution of bacterial CCMs. That is, two trajectories of gene acquisition that lead to improved growth in relatively-lower CO2 at each step. Even more surprising, neither of these pathways involved the early acquisition of a carboxysome - a 200+ megadalton protein structure that is the hallmark of bacterial CCMs. Rather, both pathways involved early acquisition of HCO3- transporters and/or carbonic anhydrase enzymes. It took us quite a while, and a fair bit of math, to understand what was going on. In short - all organisms need HCO3-. So carbon-fixing bacteria need both CO2 and HCO3-. HCO3- forms spontaneously from CO2 and water, but not very quickly. So CCM components that provide HCO3- can improve growth as environmental CO2 levels decrease over geologic timescales.
Which brings me to Reviewer #2. After the paper was accepted, this unknown scientist commented that our paper “set up straw men” only to knock them down. I truly had not intended to denigrate viewpoints we previously held by calling them “naïve,” but rather intended to walk readers through an intellectual progression that we underwent during in this project. You never know how your work is going to be received until someone else reads it, so, dearest Reviewer #2, thanks for your help. The paper is better for it.